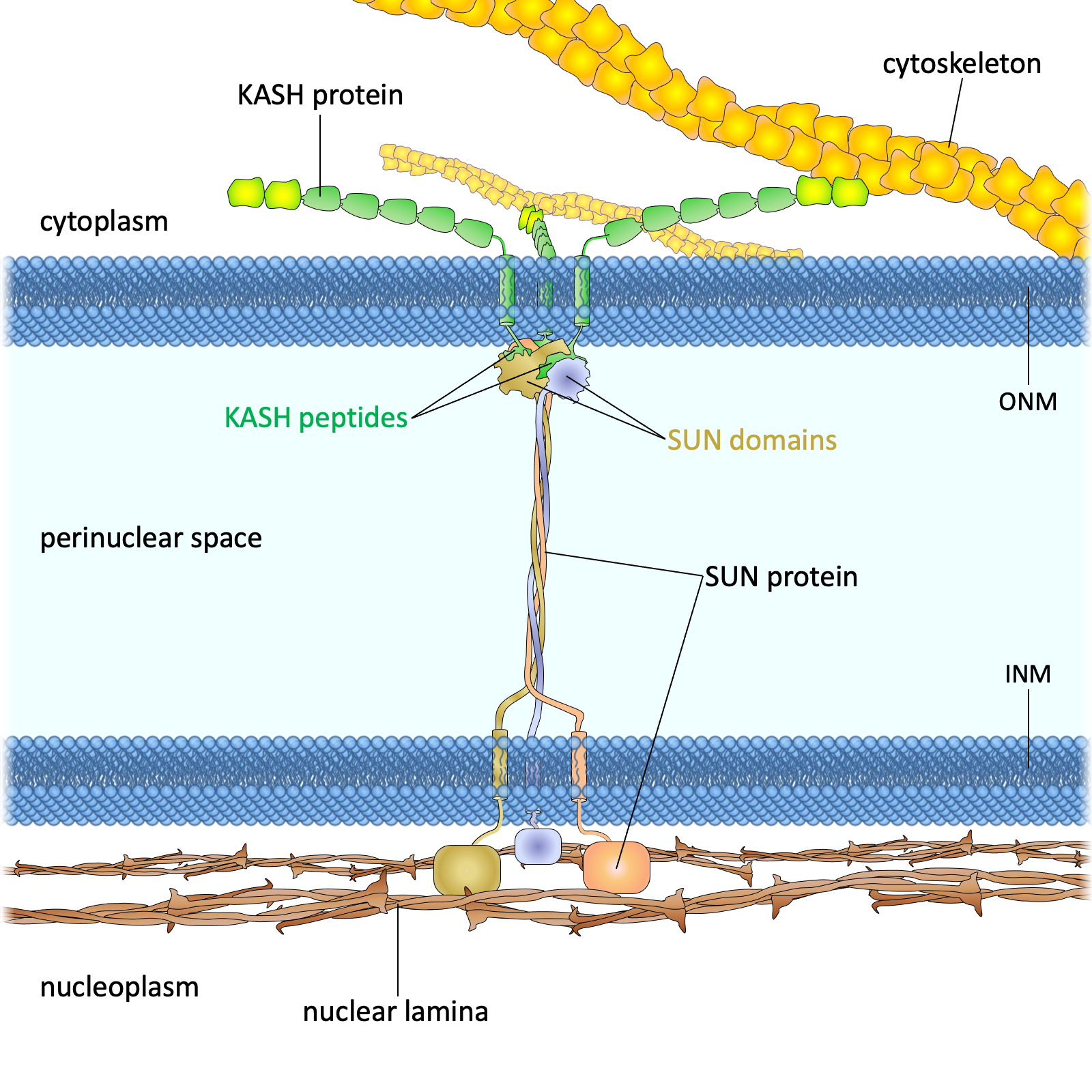
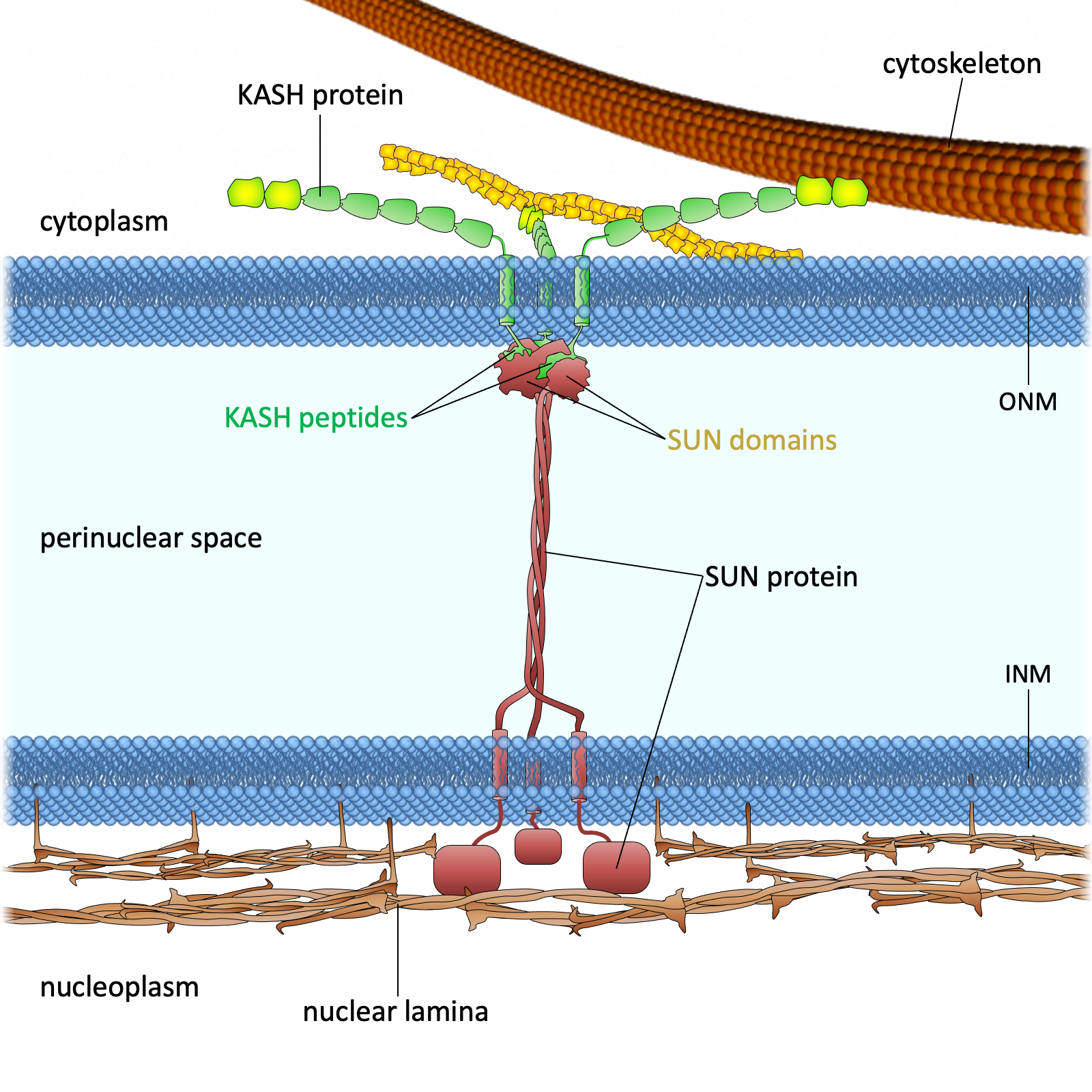
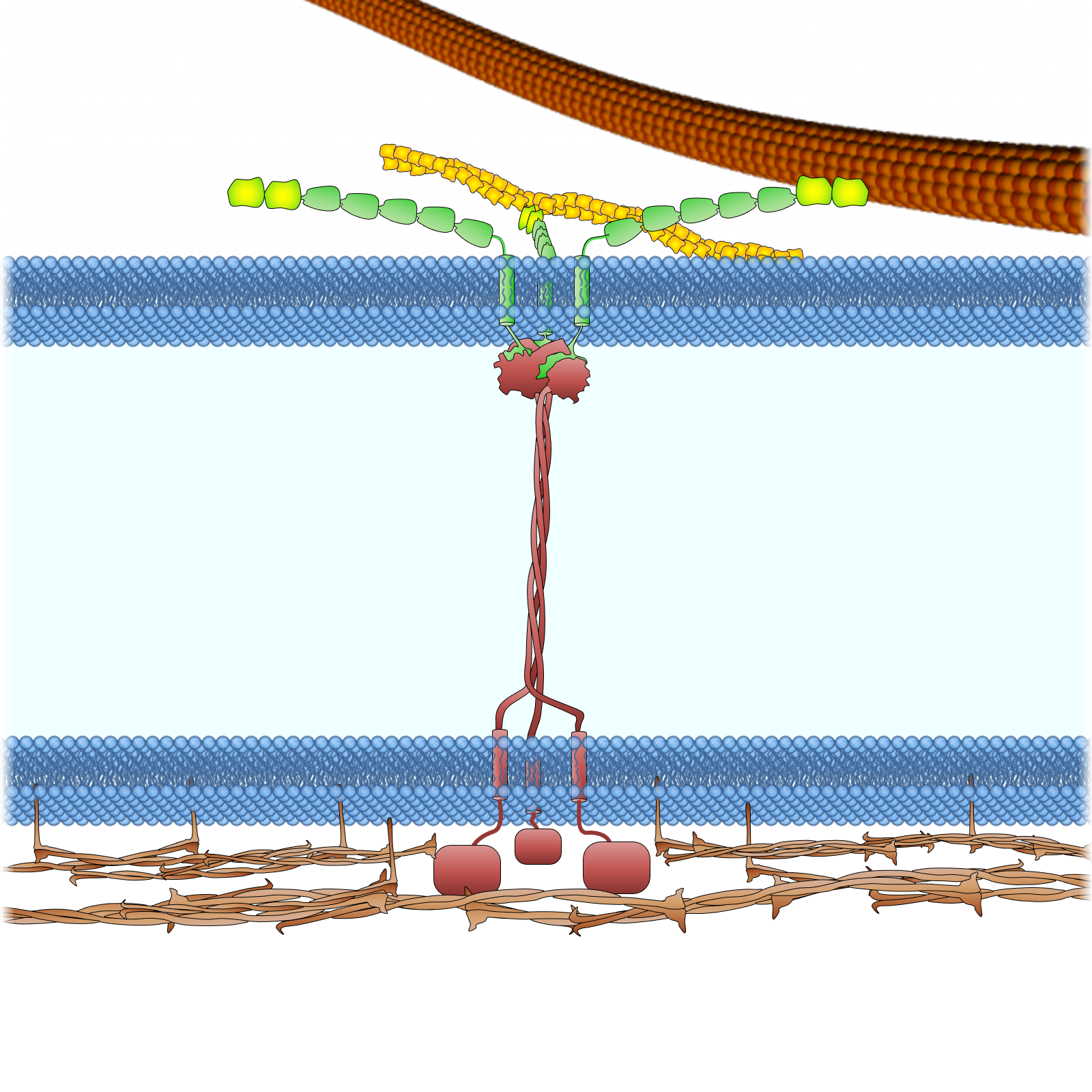
.
SARS-CoV-2 and the Nucleus.
International Journal of Biological Sciences,
2022 Jul;
18(12):4731-4743.
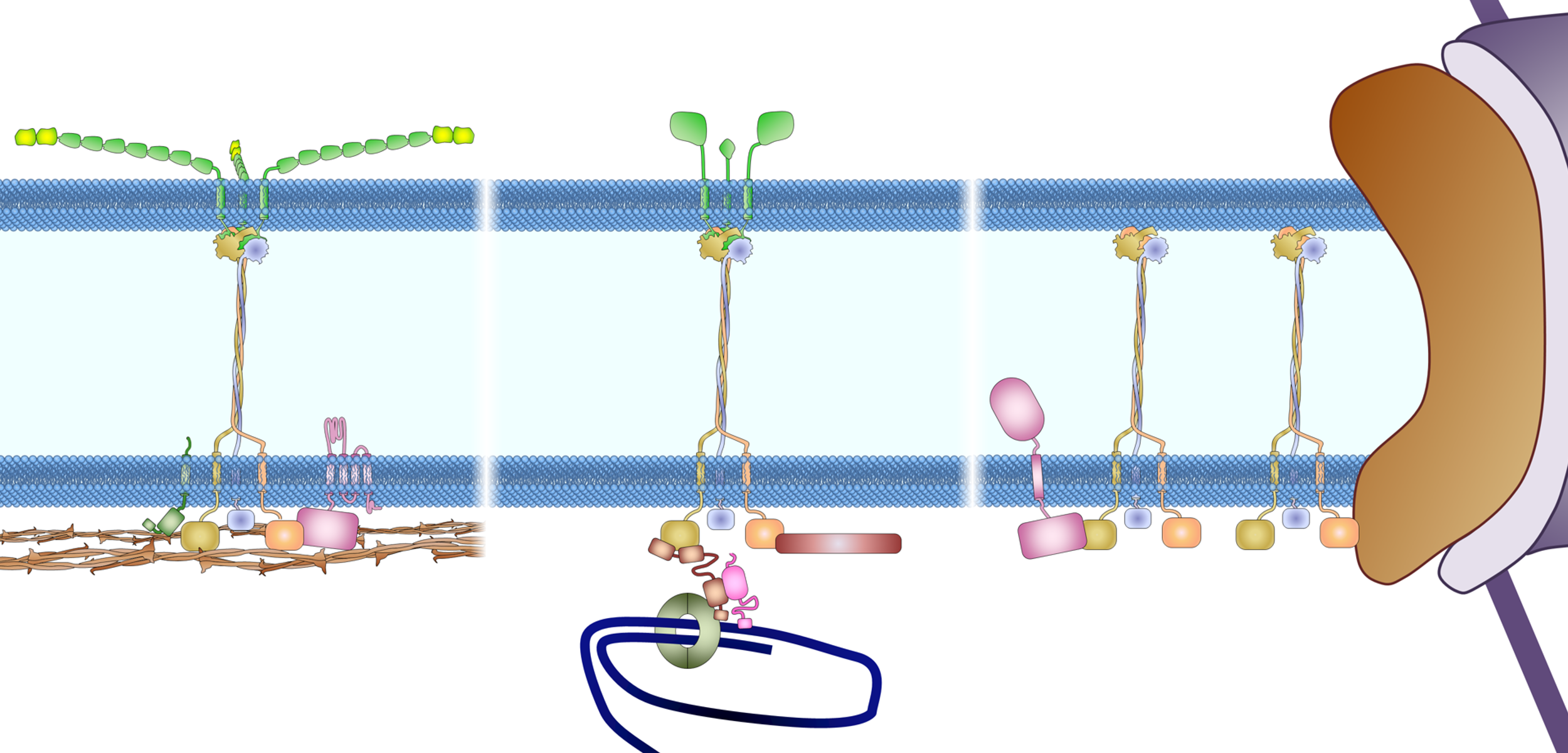
Figure 1. Nuclear import and export. Left: Cargo to be imported binds to cytoplasmic importin and enters the nucleus, where it is released from importin by Ran-GTP. The Ran-GTP/importin complex moves to the cytoplasm and binds RanBP. Ran-GAP activates the GTPase activity of Ran to convert Ran-GTP to Ran-GDP and release importin. Ran-GDP enters the nucleus and is recharged by a Ran-GEF. Right: Cargo to be exported binds to exportin and Ran-GTP in the nucleus. The complex moves to the cytoplasm where Ran-GTP is converted to Ran-GDP, leading to the disassembly of the complex. Both Ran-GDP and exportin then enter the nucleus for another round of nuclear export.
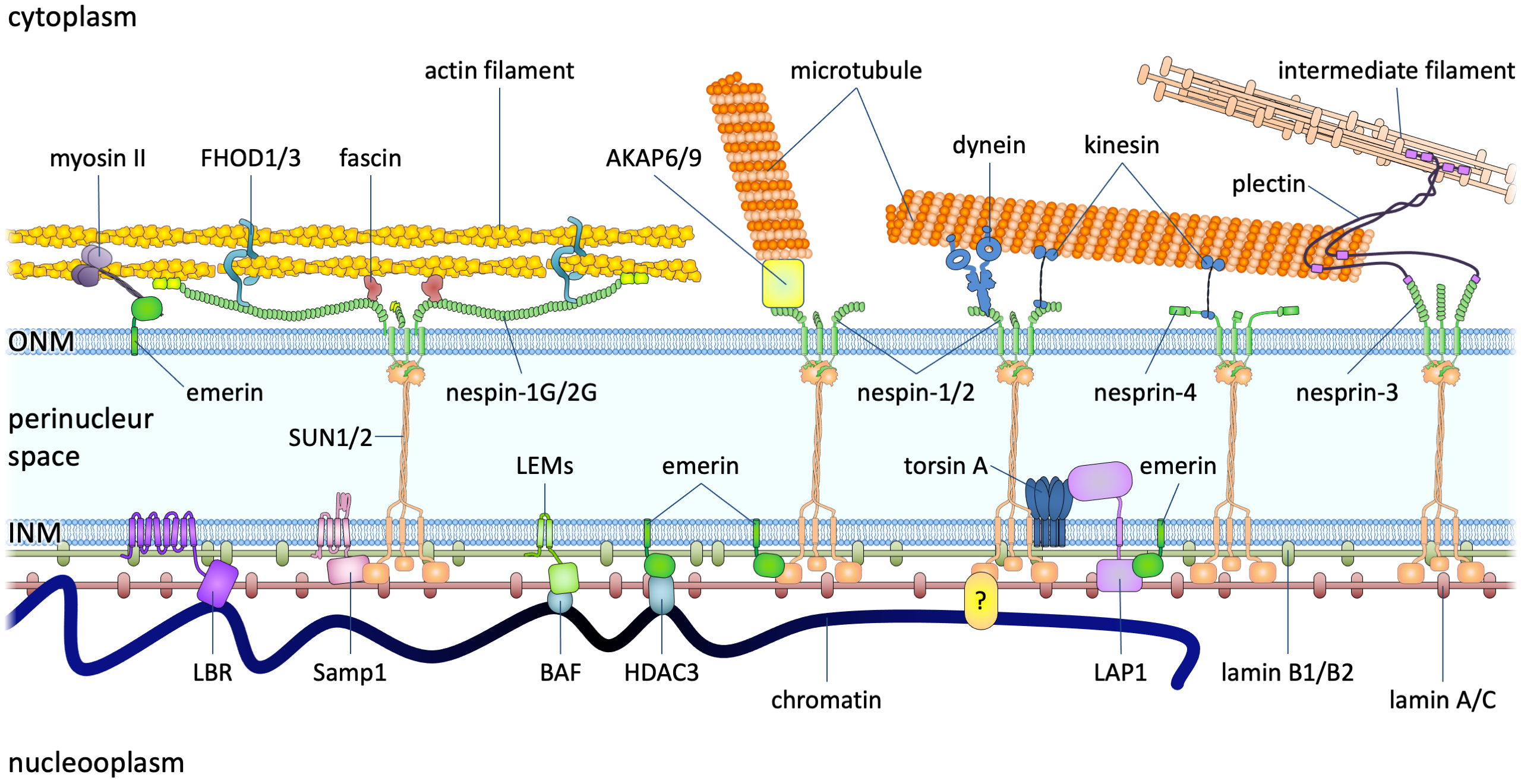
Nuclear import and export, with lables (2560 × 1440 px).

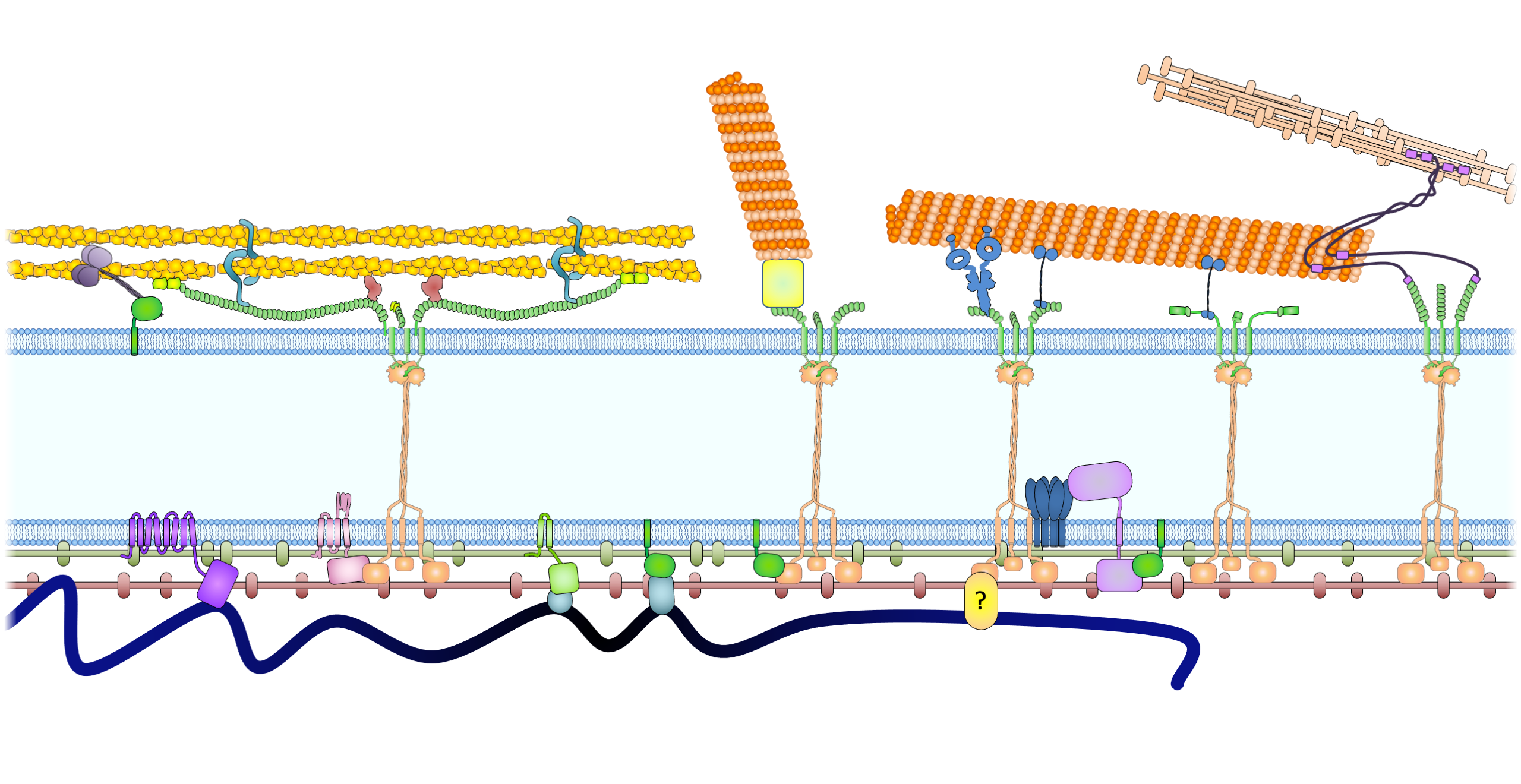
Nuclear import and export, without lables (2560 × 1440 px).
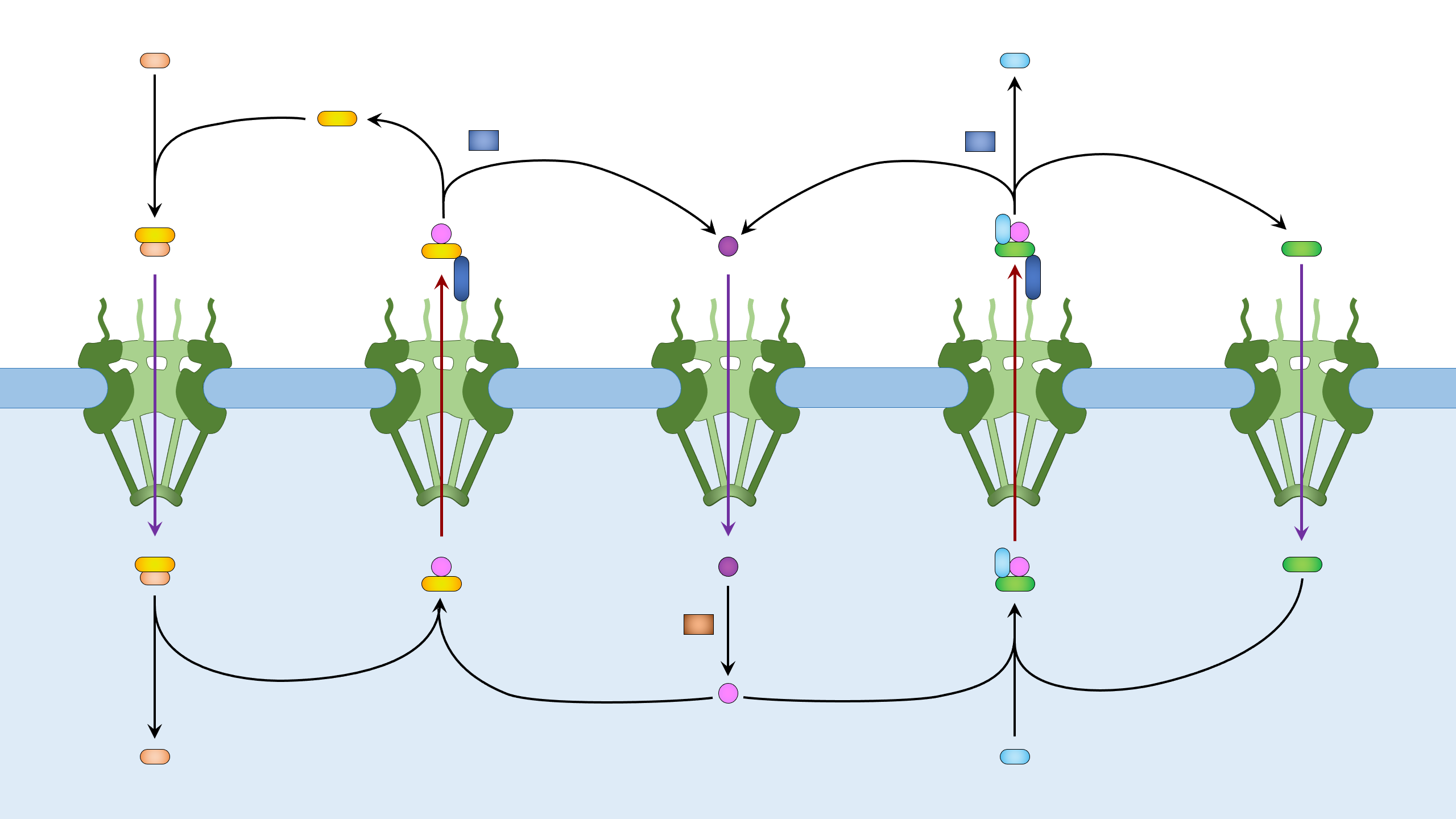
The nuclear pore complexes (2560 × 1440 px).